





The ancient araucaria family (Araucariaceae) contains three genera (Araucaria, Agathis, and Wollemia) and forty-one species of cone-bearing trees native to forested regions of the Southern Hemisphere, including South America, Malaysia, Australia, New Zealand, and New Caledonia. During the Jurassic Period, the family had an extensive distribution in both hemispheres, extending as far north as England, Greenland, and Sweden. In majestic size and beauty, araucariads rival the grander members of the pine family (Pinaceae); both families are conifers (Pinophyta). Fossil evidence indicates that ancient araucaria forests resembling present-day species date back to the age of dinosaurs. Today, araucaria forests are limited to the Southern Hemisphere and are considered a counterpart to the pine and spruce forests of the Northern Hemisphere.
Although sometimes referred to as “pines,” members of the araucaria family have seed cones and foliage that are distinctly different from those of the pine family. Trees in the pine family have cone scales with a pair of winged ovules (seeds) on the upper surface; members of the araucaria family have only one ovule per scale. The seed of Araucaria is fused to the scale and falls with the scale when the cone disintegrates. In Agathis, the seed is winged and free from the scale. Unlike the typical, slender, needlelike leaves of the pine family, the sharply pointed leaves of the araucaria family are quite variable; depending upon the species, they range from scale-like or awl-shaped to linear or oblong. Some species have broad leaves that superficially resemble the leaves of flowering plants.

Wollemia—a Surprising Discovery
Of the three genera of Araucariaceae, the most primitive is Wollemia. In 1994, David Noble discovered an unknown cone-bearing tree in the rugged Blue Mountains of Wollemi National Park, northwest of Sydney, Australia. The generic name commemorates this park and is derived from an Australian aboriginal word meaning “watch out, look around you”—an appropriate warning for anyone hiking the complex canyons and precipitous sandstone escarpments characteristic of the tree’s habitat. About forty trees in a deep narrow canyon turned out to be an undescribed species. They were named Wollemi pine (Wollemia nobilis), a remarkable “new” member of the family. Fossils resembling Wollemia and possibly related to it are widespread in Australia, New Zealand, and Antarctica, but W. nobilis is the sole living member of its genus. This rare conifer was thought to be extinct: its last fossil record was dated about two million years ago. Typical of genetic bottlenecks in small populations, chloroplast DNA studies show no discernable genetic variation among the wild trees. Like the Torrey pine (Pinus torreyi) of San Diego County, which also has little genetic variability, this is truly a relict population that was more widespread millions of years ago. The Wollemi pine is a great botanical discovery—a “living fossil.”
Dinosaur skeletons and fossils of juvenile and mature foliage of a supposedly extinct conifer have been found together throughout western Queensland in rocks dating back 120 million years. Foliage of the just-discovered Wollemi pines closely matches these fossils. Further evidence comes from live and fossilized pollen. Pollen grains of extinct plants are well preserved in ancient strata because of their durable exine coating made from a substance called sporopollenin. Although it contains only carbon, hydrogen, and oxygen, sporopollenin is one of the most stable organic compounds known. Throughout Australia and New Zealand, fossil pollen called Dilwynites has been collected from Jurassic age sediments. It has also been collected in fifty-million-year-old sediments from Antarctica’s continental shelf. Similar to araucariad pollen, it differed in its coarse, grainy coating. When compared with pollen from living Wollemi pines, there was a perfect match.


Records from paleobotanists now prove that Wollemi pines were common in moist forests on the Australian continent for countless millions of years, growing with ferns, other gymnosperms such as cycads and ginkgos, and early flowering plants. According to McLoughlin and Vajada, Wollemi pines have survived raging forest fires and the asteroid collision on Mexico’s Yucatan Peninsula that drove the dinosaurs extinct and ended the Cretaceous. Hidden away in narrow sandstone ravines for millions of years, the Wollemi pine receives the humidity and moisture necessary for its vital symbiotic mycorrhizal fungi. These habitats provided shelter from firestorms that raged in the eucalyptus forests atop the sandstone plateaus. The tree’s habit of resprouting from the base (coppicing) probably helped it recover from occasional fires within the deep, generally moist ravines.
Wollemi pines are now being propagated from both seeds and through tissue culture, and are already being cultivated in a few botanical gardens on the Pacific Coast. Time will tell if this intriguing conifer enters the commercial trade, following in the horticultural path of maidenhair tree (Ginkgo biloba) and dawn redwood (Metasequoia glyptostroboides)—both considered to be living fossils.
Wollemi pines have beautiful light green foliage composed of flattened, linear leaves. In the juvenile stage, leaves are twisted at the base to form a flattened, two-ranked arrangement similar to those of a coast redwood (Sequoia sempervirens). Adult leaves are four-ranked and spirally arranged around the stem. In their native habitat, the trees have multiple trunks; younger stems emerge from the base of the tree and gradually replace older trunks. Because of this strong coppicing habit, tree-ring dating will never reveal the actual age of an old tree. The trees are frost and shade tolerant and can also grow in full sun.

One caveat to the cultivation of this rare conifer is its susceptibility to various pathogenic soil fungi (water molds). In November 2005, wild Wollemi pines were discovered infected with a root rot fungus (Phytophthora cinnamomi); this fungus is the cause of avocado root rot, the scourge of avocado growers in San Diego County. Park rangers in Australia believe spores of this virulent water mold were introduced on the shoes of unauthorized visitors to the site; for this reason, the location of the groves has been undisclosed to the public. This vulnerability is typical of small populations with little genetic diversity. The survival of Wollemi pines in the wild could be seriously threatened, not only by such exotic diseases, but also by environmental and climatic changes.

The Araucarias
The genus Araucaria includes approximately nineteen species, all confined to the Southern Hemisphere in markedly disjunct distributions. Most of the extant araucarian taxa have evolved since early Tertiary time; extensive evolution and adaptive radiation have occurred in the islands of Australasia, where most of the species are found. Two species occur in South America and two in New Guinea (one of which also occurs in Australia), one is endemic to mainland Australia, and another is found only on Norfolk Island in the Tasman Sea. The remaining thirteen species are endemic to New Caledonia, where they occur from sea level to 3,000 feet elevation. This is a striking pattern, according to biologist Timothy Waters, because it means that this family of conifers has been successful in a tropical rain forest environment where angiosperms (flowering plants) are generally thought to have replaced conifers. He suggests that the Australasian success of araucarians may be governed more by their ability to adapt to conditions of their physical environment (including precipitation, fire, and edaphic factors), rather than their ability to compete effectively with angiosperms.
The type genus Araucaria is derived from the Arauco region of central Chile, where the Araucani Indians live. This is the land of the monkey puzzle tree (A. araucana), so named because the prickly, tangled branches would be difficult for a monkey to climb. It has been suggested that an armor of dagger-like leaves on ancient araucariads might have discouraged hungry South American herbivorous dinosaurs, such as the enormous Argentinosaurus that weighed an estimated eighty to a hundred tons! Monkey puzzle trees do not grow well in Southern California, but are occasionally planted in parks and gardens of Northern California. They grow much better in the colder, wetter climates of Oregon, Washington, and British Columbia. Another ancient South American species called pino paraná or paraná pine (A. angustifolia), native to southern Brazil and Argentina, is occasionally grown in coastal California.
Australian members of the Araucariaceae commonly grown in California include the tall, prickly-leaved bunya-bunya (A. bidwillii) of Queensland, with huge pineapple-shaped cones, hoop pine (A. cunninghamii), an important Australian timber tree, and Norfolk Island pine (A. heterophylla), native to Norfolk Island. The latter species is commonly grown in parks and gardens in the San Francisco Bay Area, and in containers in Southern California; it is often sold as star pine, because of its horizontal tiers of radiating branches. Norfolk Island pine was discovered by Captain James Cook on his second voyage to Australia and New Zealand aboard the HMS Resolution (1772-1775). Bunyabunya also has an unmistakable silhouette, with barren, horizontal limbs tufted at the ends with spiny leaves. Its huge seed cones pose aserious threat to unsuspecting persons standing beneath the canopy. Unlike most conifer species, wild populations of Araucaria are typically dioecious, with pollen-bearing and seedbearing cones on separate male and female trees. Trees are occasionally monoecious, and there are remarkable but unsubstantiated reports of trees that change sex with time.
In Southern California, there are also trees that resemble Norfolk Island pines, but with more closely spaced limbs and a narrow, columnar, crown that resemble Cook pines (Araucaria columnaris). According to Angela Keplar’s Trees of Hawaii, most of the trees called Norfolk Island pine in Hawaii are actually Cook pines. They have naturalized throughout the Hawaiian Islands and have been exported as lumber and container-grown “Christmas trees” to the US mainland. Both species were introduced to Hawaii in the late 1800s. Some botanists consider the narrow-growing trees in Southern California to be hybrids of the two species, while others insist they are the true Cook pine.
In general, the interval between tiers of limbs is greater on Norfolk Island pines, particularly on younger trees, and the horizontal limbs are longer than those of Cook pines; the latter has a slender, spire-like crown with shorter, more closely spaced horizontal branches. When the two species are compared side-by-side, the differences between them are striking; however, there are also trees with intermediate branching patterns. Leaf shape and size is variable and not particularly useful in separating these two species. Although A. columnaris is not listed in the latest Sunset Western Garden Book, there are confirmed specimens of this tree in Southern California at Quail Botanical Gardens, Huntington Botanical Gardens, and the Palomar College Arboretum.
Phylogenetic studies comparing chloroplast DNA have shown that the Norfolk Island pine’s closest relatives are a group of thirteen species of Araucaria endemic to New Caledonia, including Cook pine. Although New Caledonia is a relatively small island of only 19,000 square kilometers, it contains a rich conifer flora of forty-three endemic species, including two-thirds of the world’s Araucaria species and five of the twenty-one species of Agathis. The New Caledonian species are seldom cultivated in California, except by the occasional passionate collector.

The Resinous Agathis
The genus Agathis includes thirteen species of large, resinous, broad-leaved conifers scattered throughout Australia, New Zealand, and the Malay Archipelago. Several species are the source of timber and valuable copal varnish, including kauri pine (A. australis) of New Zealand, Amboina pine (A. dammara) of Malaysia, and Queensland kauri (A. robusta) of eastern Australia. One of the largest kauri pines found on New Zealand’s North Island was documented at 169 feet tall with a trunk forty-five feet in circumference; it is thought to be a little more than 2,000 years old. The Maori name for this individual is Tane Mahuta, which means “god of the forest.”
Kauri pines and Queensland kauri are occasionally found in the collections of various botanical gardens in both Northern and Southern California. Their extremely slow growth rate and ultimate great size may limit their use in private gardens. Both are notable for their relatively smooth trunks with bark that exfoliates in irregular patches.
Copals are a group of resins that form particularly hard varnishes. Dammars are another group of hard, durable varnishes that turn shiny and transparent when dry. Although some species of Agathis are named dammar, most dammar resins come from tropical Asian trees of the genus Shorea in the Diptocarpaceae. Copal and dammar resins improve the drying qualities and provide the luminous depth and brilliance for which oil paintings are known.

The Fossil Story
Fossil evidence indicates that the aracauria family reached its maximum diversity during the Jurassic and Cretaceous periods, between 200 and 65 million years ago, with worldwide distributions. At the end of the Cretaceous, when dinosaurs became extinct, so did the Araucariaceae in the Northern Hemisphere. Until about 135 million years ago, trees of the Araucariaceae grew in forests of the ancient southern supercontinent called Gondwana, which combined the land masses now known as South America, Africa, Antarctica, India, and Australia. By sixty-five million years ago, the continents had drifted into positions resembling their present-day configuration.
Petrified Forest National Park in Arizona protects hundreds of acres of perfectly preserved logs from an ancient conifer forest dating back to the late Triassic Period (approximately 225 million years ago). Streams carried fallen logs into this once swampy lowland region where they were buried in sediments rich in volcanic ash. Over countless centuries, the woody tissue of the logs was replaced with minerals and gradually turned into stone. Many of the reddish, agatized logs do not show any cellular detail; however, there are some permineralized specimens in which minerals permeated the porous cell walls and filled the cell cavities (lumens). Thin sections of these samples, when viewed under a microscope, reveal remarkable cellular detail.
The trees of this extinct forest coexisted with dinosaurs. Most of the petrified logs were previously assigned to the extinct Araucarioxylon arizonicum, a presumed distant relative of Araucaria. Although that binomial has been used in botanical literature for more than a century, Rodney A Savidge of the University of New Brunswick has concluded that it is superfluous and therefore an illegitimate name. He examined thin sections of the original three specimens housed at the Smithsonian Institution upon which the species was first described by FH Knowlton in 1889 and found that they represented three different species within two new genera of extinct trees. Only one of the three specimens was retained as the new type Pullisilvaxylon arizonicum. Savidge examined several other logs previously identified as A. arizonicum and concluded that they represented additional new genera and species. His extensive anatomical studies indicate that the majority of logs in the Petrified Forest National Park do not belong to a single species but rather to a complex of extinct conifers. Based solely on the xylem structure of permineralized wood (including resin canals, rays, and tracheid pitting), and without seed cones or DNA evidence, it is difficult to be certain which trees (if any) in this complex are ancestral relatives of the araucaria family.
Trees in this diverse forest grew to a height of 200 feet with a trunk diameter of from four to nine feet. According to paleobotanists Sidney R Ash and Geoffrey T Creber, the living trees did not closely resemble any of the present-day Araucaria species of the southern hemisphere. The branches did not occur in whorls as they do in most conifers but grew irregularly along the trunk, nor did the bark resemble that of living species. These ancient trees flourished during a time when all of the continents were united into the vast supercontinent Pangaea, which broke up in the middle of the Jurassic period. The area of Petrified National Park was, at that time, located near the equator, at approximately the latitude of present-day Central America. The trees grew in a tropical rain forest with marshes, rivers, and lakes—an environment totally unlike today’s Arizona landscape.
Another rich fossil area for Jurassic age araucarian forests is Cerro Cuadrado Petrified Forest National Monument in Patagonia, Argentina. Some of the largest logs in this arid desert region are ten feet in diameter, the remnants of trees that were over 200 feet tall. Seed cones of Araucaria mirabilis from this site are remarkably preserved in every detail. The cones clearly show one seed per scale and resemble modern cones of Norfolk Island and Cook pines. The oldest documented record for resin canals comes from cone scales of this extinct conifer.

Resins: Beautiful and Useful
Amber is ancient plant resin that has metamorphosed into a hard, plastic-like polymer over millions of years. Insects often become trapped in the sticky sap and are perfectly preserved in a transparent tomb of fossilized resin. Baltic amber dates back to the early Tertiary Period, approximately fifty million years ago. Ninety percent of this amber appears to be from a single plant source. For decades, Baltic amber (succinite) had been arbitrarily assigned to an extinct pine (Pinus succinifera) because of the presence of succinic acid; however, infrared spectroscopy studies now show that Baltic amber may be more closely related to resins of broad-leaved conifers of the Araucariaceae. According to biologist Jean H Langenheim, an authority on resins, Baltic amber contains pinaceous inclusions (wood fragments and cones) but with araucarian chemical characteristics, so the origin of these vast deposits remains an enigma.
In New Zealand, living araucariad forests of kauri pine produce copious amounts of resin that once formed a thriving industry for hard, durable varnishes and linoleum. Large lumps of hardened resin (up to one hundred pounds in size) were dug out of the ground in extensive forested areas of the North Island. Forests such as this may have once flourished in the Baltic region sixty million years ago. Throughout the world, the most copious resin-producing trees occur in tropical regions. These complex mixtures of terpene resins may have evolved to serve as a chemical defense against the high diversity of plant-eating insects and parasitic fungi found in the tropics.
Relevant today for their practical uses and their distinctive character in the landscape, members of the araucaria family resonate for us through time. A walk along shaded pathways lined with araucariads reveals a view deep into the geologic past. Having changed little during the past 180 million years, these living fossils are resilient, successful, cone-bearing trees that link us to a time when dinosaurs ruled the earth.

Bibliography:
Armstrong, Wayne P. 2008. “Cycad Distribution and Continental Drift.” Available online at: https://waynesword.palomar.edu/cntdrift.htm.
Ash, Sidney R, and Geoffrey T Creber. 2003. “The Late Triassic Araucarioxylon arizonicum Trees of the Petrified Forest National Park, Arizona, USA.” Paleontology 43 (1): 15-28.
Bitner, Richard L. 2007. Conifers for Gardens: An Illustrated Encyclopedia. Portland, Oregon: Timber Press.
Farjon, Aljos. 2008. A Natural History of Conifers. Portland, Oregon: Timber Press.
Kershaw, Peter, and Barbara Wagstaff. 2001. “The Southern Conifer Family Araucariaceae: History, Status, and Value for Paleoenvironmental Reconstruction.” Annual Review of Ecology and Systematics 32: 397-414.
Langenheim, Jean H. 2003. Plant Resins: Chemistry, Evolution, Ecology, and Ethnobotany. Portland, Oregon: Timber Press.
McLoughlin, Stephen, and Vivi Vajda. 2005. “Ancient Wollemi Pines Resurgent.” American Scientist 93: 540-547.
Savidge, Rodney A. 2007. “Wood Anatomy of Late Triassic Trees in Petrified Forest National Park, Arizona, USA, in Relation to Araucarioxylon arzonicum Knowlton, 1889.” Bulletin of Geosciences 82 (4): 301-328.
Waters, Timothy, Chloé A Galley, Russell Palmer, Samuel T Turvey, and Nicholas M Wilkinson. 2004. “Report of the Oxford University Expedition to New Caledonia, December 2000 –January 2001.”
Woodford, James. The Wollemi Pine. 2000. Melbourne, Australia: The Text Publishing Company.





Ground Up Science for Greener Cities with Garden Futurist Dr. Alessandro Ossola
Spring 2023 Listen to the Podcast here. Alessandro Ossola is a scientist who gets very excited about the challenge of climate change allowing for an

Readying Urban Forests for Climate Realities with Garden Futurist Dr. Greg McPherson
Winter 2023 Listen to the Podcast here. “Going from the mow and blow to a more horticulturally knowledgeable approach to maintaining the landscape. And that
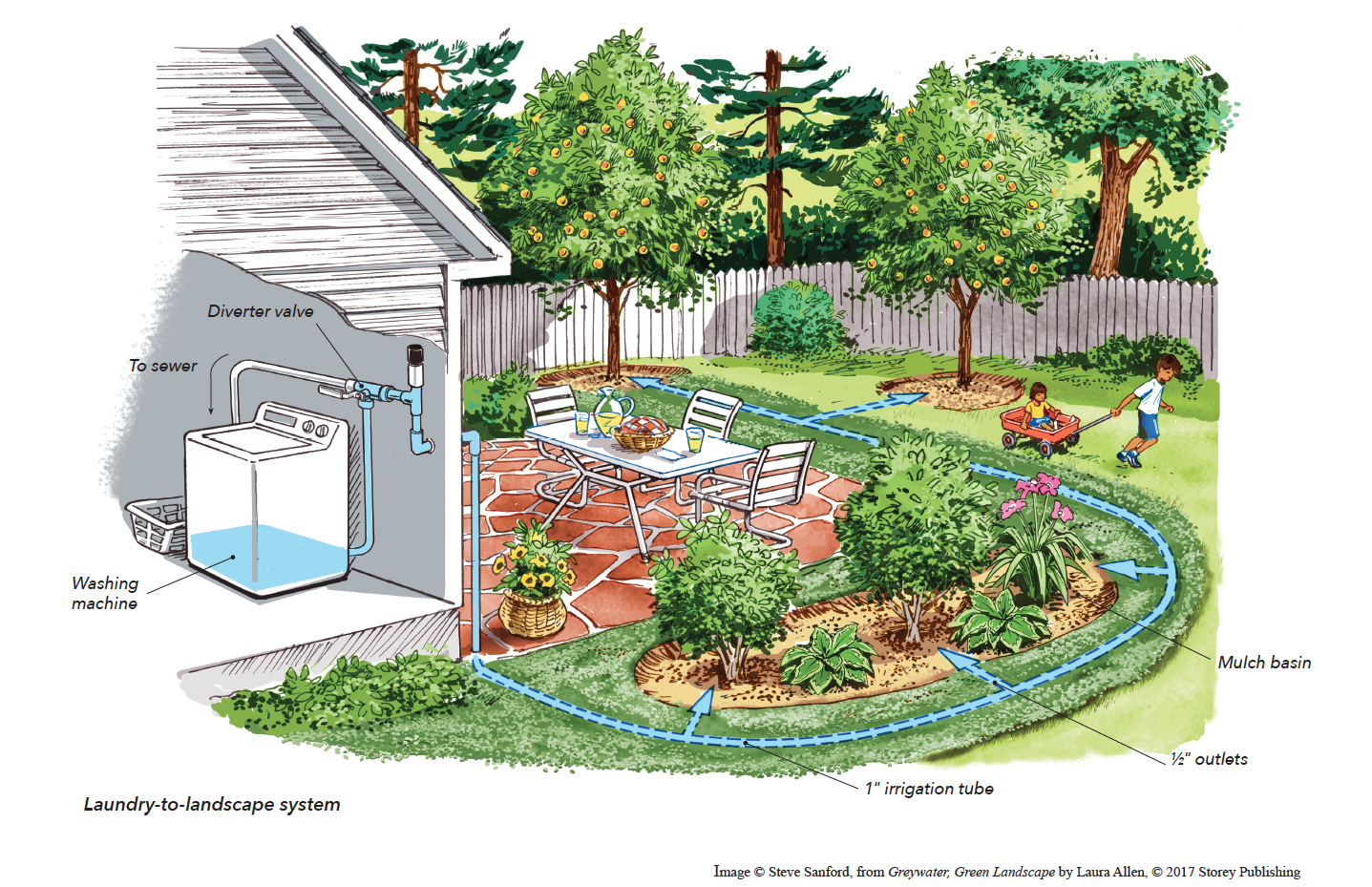
Welcome, Greywater, to the Garden
Summer 2022 Oh, summer: delightful warm air, tomatoes swelling on the vine, fragrant blooms on an evening stroll. When it’s warm and rainless, how is

Big Tree-Data and Big-Tree Data with Garden Futurist Matt Ritter
Summer 2022 Listen to the full Garden Futurist: Episode XV podcast here. We are in an environmental crisis right now in many parts of California
Responses