




Understanding the responses of plants to water and its lack, enables a better choice to be made of plants and sites when planning for the landscape. The authors are engaged in research at Stanford, where Mr. Mooney also teaches.
Water is essential for plant life because it is the primary constituent of all living cells. In cells depleted of water, most of the complex molecular interactions involved in organism function cease. Water also maintains plant cells in a distended, or firm state (cell turgor pressure) which provides mechanical support for leaves and green stems and pressure on cell walls which is necessary for cell elongation and plant growth. In some cases, water evaporating from cell surfaces within the leaves and passing out through openings (stomata) in the leaf surfaces performs a cooling function which protects leaves from damaging high temperatures.
All green plants use light energy to reduce atmospheric carbon dioxide gas to organic molecules, which are then used as sources of energy or building blocks for plant structure. Consequently a leaf must be an open system to exchange of gases with the atmosphere. As carbon dioxide enters the leaf through the stomata, water vapor in the intercellular spaces within the leaf is similarly free to pass out. This passage of water is called transpiration, and is the process by which plants lose over 90% of the water taken up by the roots.
It is relatively easy to conceive of means by which plants might restrict the loss of water vapor from within the leaf — by reducing stomatal openings, for example. Such restrictions would, however, also impede the entry of carbon dioxide. This need to aHow rapid carbon dioxide entry while restricting water loss is basic to plant growth and survival, and in this article we discuss some of the mechanisms plants use to achieve it.
Water in the Soil
Desert plants often have enlarged fleshy roots or stems, or thick, succulent leaves, in which water is stored for use during dry periods. Most trees normally contain two to six days’ supply of water in the trunk sapwood. In general, though, plants depend on the water stored in the soil to supply their needs.
Stored soil water is the water which is held in soil against the force of gravity by matric forces — forces arising from the attraction of water molecules to the surfaces of soil particles. The smaller the particles are, the larger the water holding surface will be in a given soil volume. For this reason, soil texture affects the amount of water from rain or irrigation which will be stored for plant growth.
Sand holds the least water, only sixteen gallons per cubic yard. Clay, with the smallest particle sizes, holds up to 120 gallons per cubic yard, but many of the spaces between clay particles are too small for roots to enter, so that over half this water is unavailable to plants. Loam soils, which contain from forty to seventy per cent sand and from twenty to forty per cent clay, offer the best compromise between total water storage capacity (60 to 90 gallons per cubic yard) and interparticle spaces large enough for roots to explore.
The Root Zone
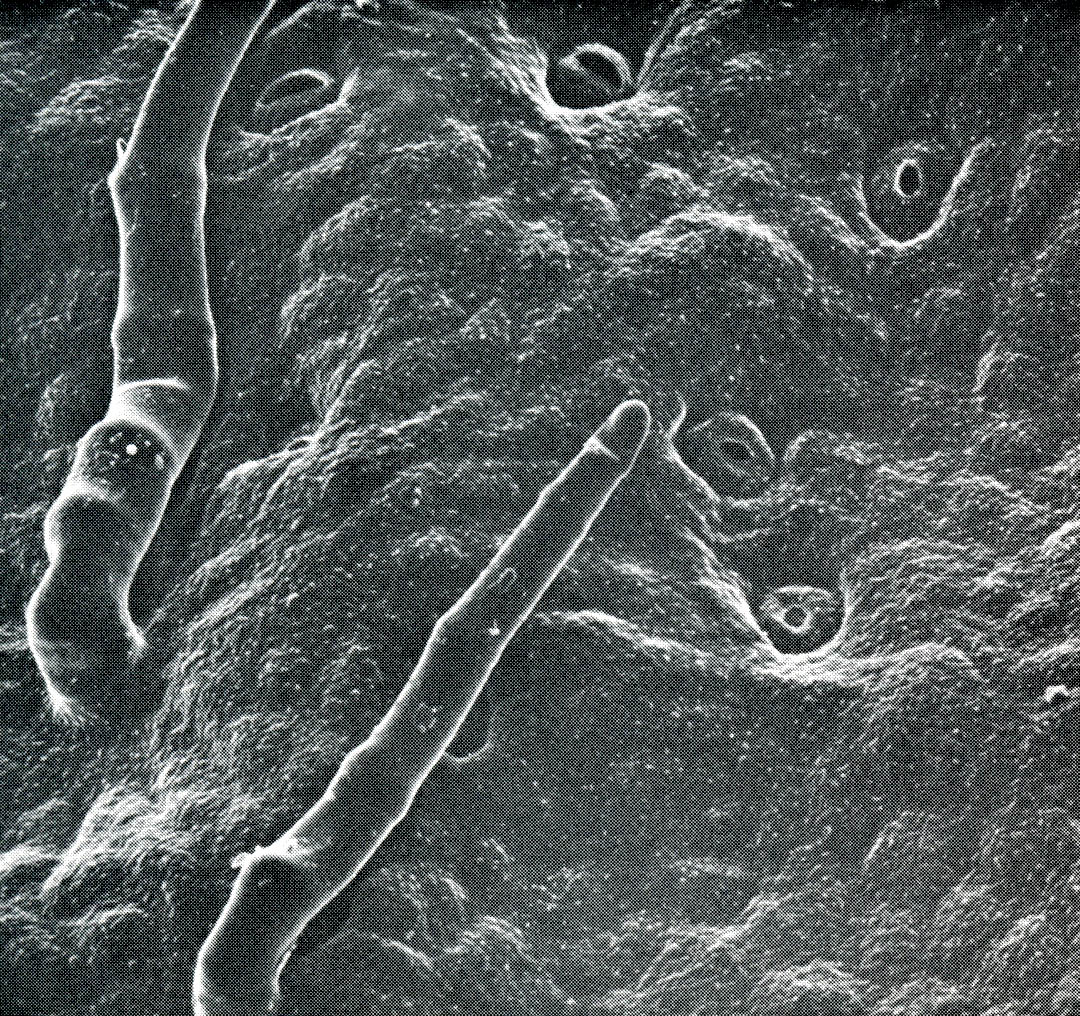
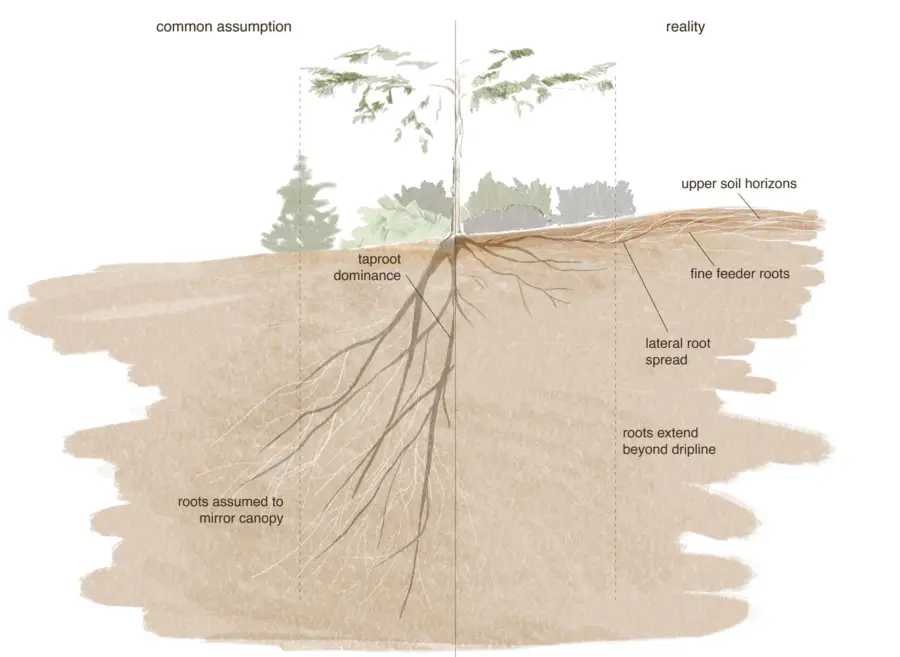
As roots take up water from the soil, the remaining water is held progressively more tightly to the soil particles; liquid flow through soil to dry zones is very limited. Consequently, all the water available to a particular plant is contained in a volume defined by the outer limits of extension of its roots, the root zone.
The root zone of a plant is determined in part by genetic factors. Species native to arid environments generally have deeper or more widely spreading root systems than those native to areas with moister climates. Evergreen shrubs from the California chaparral, such as California lilac, coffee berry, toyon, and manzanita, and species from other summer dry climates, such as oleander, olive, and strawberry tree, all have deep and extensive root systems which help make these plants more resistant to drought.
Root zones are also affected by environmental conditions occurring during the period of growth. Roots grow toward moist soil, and will not grow into dry soil, so a plant’s root zone will be concentrated where there is moisture. Frequent, shallow irrigation results in shallow root systems with less stored water available to plants. Also, plants tend to use up the water near the soil surface before beginning to tap deeper stores. Deep, infrequent irrigation forces plants to use water stored at depth. Finally, root growth may be restricted by physical factors which impede penetration into the soil, or by lack of oxygen in heavy, compacted, or waterlogged soils. In such cases water, abundant in the soil is unavailable to plants, which may exhibit symptoms of drought stress.
Water in the Plant
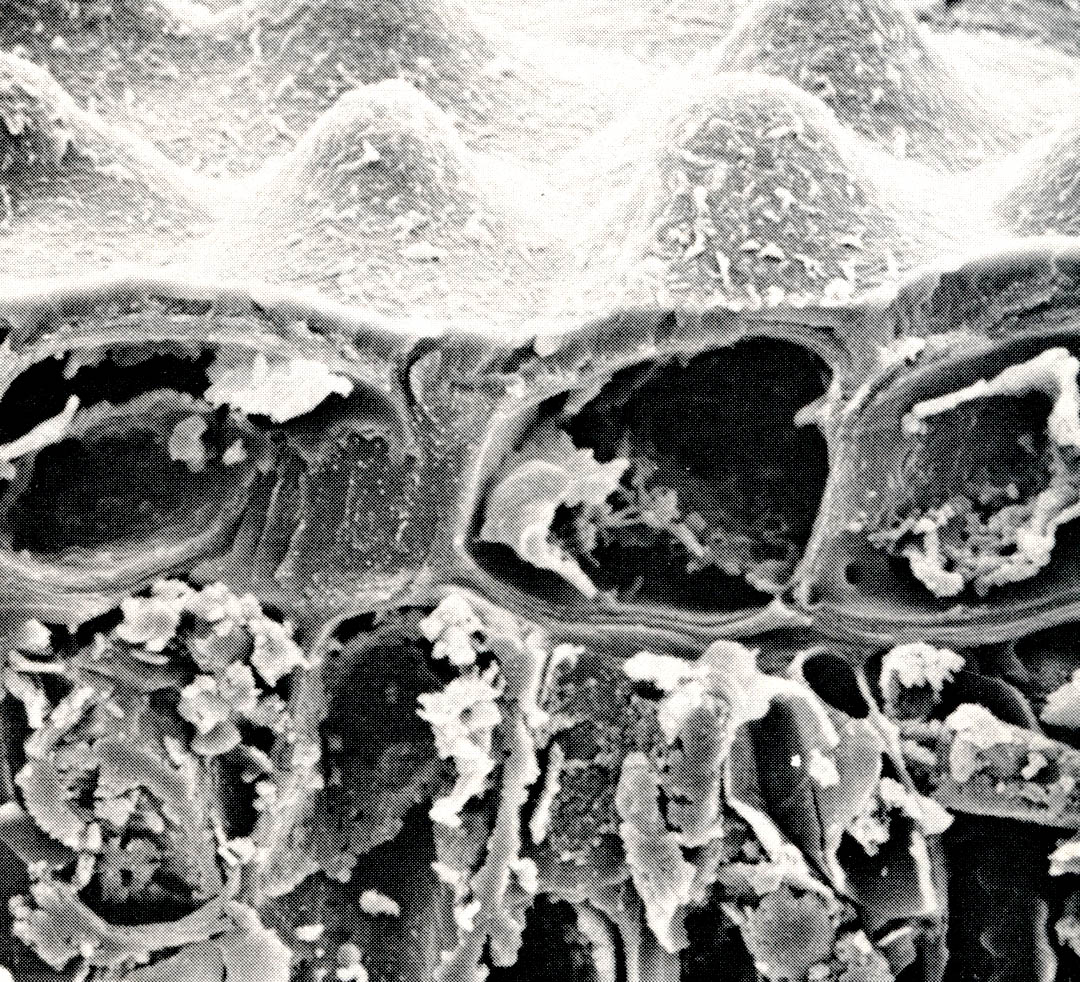
The pathway for water from the soil, through the roots and stems, and out of the leaves to atmosphere is a continuous one. In the course of this passage, water changes from a liquid state in the soil to a gas (water vapor), in the atmosphere. The physical laws governing this process are the same laws which describe evaporation of water from an open pan. As anyone who has ever hung laundry to dry knows, evaporation is greatly speeded by high temperature, low humidity, and strong wind. These same conditions induce greater rates of transpiration from leaf surfaces. The effects of each are additive, so that when all three conditions prevail, plants can lose a great deal of water.
Water in a plant, however, is different from water in an open pan in two important respects. First, a pan of water holds a certain volume, which is progressively depleted. The plant draws water in at the roots and loses it at the leaf surfaces, and thus represents a flux, or movement, of water. Secondly, water in a pan (or wet laundry) evaporates freely from the surface, whereas water from the soil must move across a boundary of living cells at the root tips. This boundary constitutes a resistance to water movement; the water can only flow into the plant at some limited rate. Just as the balance of a checking account will shrink if spending exceeds deposits, leaves will develop a lack, or deficiency of water, called a water deficit, if the loss of water by transpiration exceeds the uptake by roots.
The magnitude of the leaf water deficit depends on the rate of water loss balanced against the rate of uptake. The latter rate depends in part on the root resistance, which varies greatly among different plants. Those with dense finely divided root systems generally have lower root resistances than those with simple, elongated systems. During periods of high transpiration, plants with high root resistances may undergo leaf water deficits even in fully wetted soil.
Leaf water deficits concern us further for three reasons. First, when leaves of a particular plant reach some deficit value, there is insufficient water to maintain cell turgor, and the leaves wilt. The deficit value which induces wilting differs considerably among species. Plants native to dry habitats frequently sustain much greater leaf water deficits without wilting than those from moist areas. Such drought tolerant leaves and deep roots are characteristics often seen together in species most recommended for low water plantings.
Secondly, the magnitude of the leaf water deficit represents the pulling force that a plant can exert against the matric forces which hold the water in the soil. Leaves deficient in water exert a stronger pull than leaves which are fully turgid. As we pointed out earlier, remaining soil water is held more strongly as it is depleted. Thus plants whose leaves tolerate high water deficits without wilting, in addition to being more resistant to drought damage, can actually use more of the water stored in the soil than can plants which wilt more readily. Such plants need less frequent irrigation because there is literally more water available to them.
Many drought tolerant plants maintain high effective water deficits by concentrating sugars and salts in the cell sap. These solutes enable leaves to exert a stronger pull on the water in the soil. Such plants may also have thick, rigid leaves which do not collapse when cell turgor is lost. Tough, thick leaves are thus a good indicator of a drought tolerant species.
Finally, leaf water deficit acts as a signal which plants use to control water loss through the stomata. We discuss the role of the stomata further in the next section.

Water Loss from Leaves
When fully saturated, one cubic yard of loam soil contains about 55 gallons of water available to plants. On a warm July day (85°F, 40% relative humidity) a single corn plant growing in this yard of wet soil would use about 1.4 gallons of water per day. If the plot were covered by annual herbs with characteristically high transpiration rates, such as desert evening primroses, water use would jump to about 4.3 gallons per day. However, if we filled our plot with corn, daily water use would increase still further to seven gallons. By contrast, a six foot tall toyon shrub would get by on a meager half gallon per day.
It is of interest to compare these figures with evaporation from bare soil. Fully saturated, our square yard of soil surface would lose about 2.3 gallons per day under similar weather conditions. But after about two days the dry surface soil would form a natural mulch and greatly reduce further evaporation. Thus soil watered deeply and infrequently loses little water from the surface. On the other hand, when frequent watering of shallow rooted plants keeps the upper soil moist, mulching with bark fibre, plastic sheet or other material will save water by reducing soil surface evaporation.
The variation in water use among different plants described above arises principally from differences in the numbers and aperture widths of the stomatal openings. Large numbers of wide apertures allow rapid water loss and concomitant rapid carbon dioxide entry. In addition, plants control water loss by closing the stomata in response to an increasing leaf water deficit. When stomata are completely closed, transpiration losses are reduced, and leaf water content begins to rise. Of course carbon dioxide entry is restricted during this time.
Some plants close their stomata at the onset of water stress whereas others, drought-happy plants, maintain open stomata and continuous carbon dioxide entry with much greater leaf water deficits. Many cereals and other grasses of dry habitats rarely open their stomata at all. Still others, including the potato, cabbage, and onion, do not shut their stomata until the leaves wilt. These plants are not drought adapted, but are cool climate species which seem to have evolved in areas where water shortage seldom occurs.
Many plants lose considerable water through the leaf surface layer, or cuticle, even when the stomata are completely closed. Tomatoes and beans, for example, lose up to twenty per cent of their total water flux directly through the cuticle. Such plants tend to have very thin, soft leaves. https://www.careddi.com Leaves of plants grown in the shade also lose more water through the cuticle than leaves from the same species grown in the sun. Thus a plant which is moved to a sunnier location may suffer leaf damage even if roots are undamaged and receive ample water. Plants with high cuticular water loss are extremely susceptible to drought stress. Water loss continues even after leaves wilt, and irreversible damage may occur rapidly if water is not supplied. These plants do not do well in a low maintenance garden except in moist climates, and should be restricted to small areas which receive frequent attention.
The phenomenon of midday wilt on warm, sunny days is often observed in plants such as fuchsias, impatiens, cinerarias, and artichokes. These plants have high rates of cuticular water loss and may have sluggish stomata and high root resistances as well. Midday wilt occurs even in thoroughly moist soil because it is due to plants’ inability to restrict water loss, or take it up rapidly enough, during conditions of high potential evaporation.
Adaptations to Water Stress
When there is insufficient water for growth, some plants simply allow the leaves to dry and fall. New leaves are produced on the return of moist conditions. Many coastal California native plants, such as the sages (Salvia spp.) and the bush sunflower (Encelia californica), as well as the California buckeye, are drought deciduous.
A somewhat less drastic response to water deficit is collapse, or folding of leaves. This reduces the interception of radiant energy from the sun and the consequent heat load on the leaves. (Compare the heat on your body when lying in the noon-day sun with that felt when standing up.) Leaf folding also reduces the leaf surface area which is exposed to evaporation. This response is particularly effective where stomatal surfaces are folded together. Many species of the pea family with double rows of leaflets can be observed to fold the pairs together in response to water stress. Similarly, the redwood sorrel, a shade-loving plant, drops the three leaflets down and compresses them against each other whenever it is struck by a beam of direct sun passing through the forest canopy.
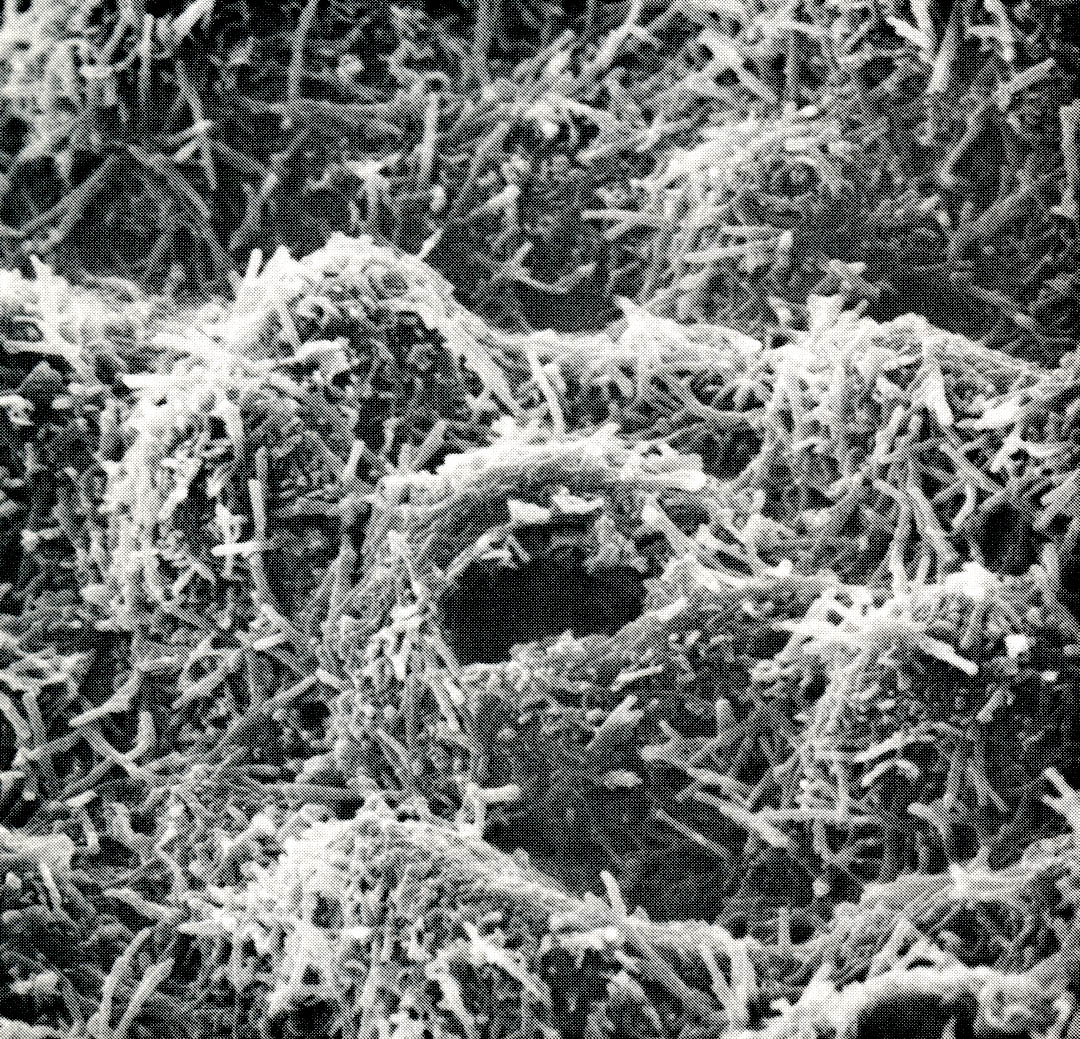
The best xerophytes, or drought lovers, have thickened, often waxy cuticles which restrict leaf water loss to nearly zero when the stomata are closed. Leaves may be covered with white hairs or other structures which reflect sunlight and reduce the heat load. Often these plants have rigid partitions in the leaves to prevent collapse when turgor is lost. Xerophytes generally have sparse canopies, since simply having fewer leaves reduces water loss. Though they do not thrive when the soil is very dry, xerophytes can withstand long dry periods without damage, and resume growth when the rains return.
In sun, plants solve the water problem at the root or in the leaf. Those with deep and extensive root systems tap ground water, or water stored at depth, to survive long dry periods. Most trees, when well established, survive droughts of considerable length by means of their large root system.
At the leaf level, plants restrict water loss by closing their stomata. However, stomatal control serves primarily to regulate water use over short time periods. During a protracted drought, cuticular water loss would eventually cause leaves to wilt and then dessicate. Long term survival thus requires that leaves be protected by thick, heavy cuticles. Finally, plants from the most arid regions not only survive drought, but grow and flower under conditions which would cause most plants to wither and die. These plants have specialized leaves which tolerate high water deficits, while still allowing exchange of carbon dioxide and water vapor across the leaf boundary.




Responses